Davidia involucrata, endemic to central China (Ying et al. 1993), is a deciduous tree up to 20 meters tall, popular in botanical gardens of Europe, North America and Australia as well as Asia (Sun & Huang, 2011). Known as the dove tree, or handkerchief tree because of its showy, white, inflorescence bracts (Fig. 1), it is related to the dogwoods, placed in the asterid angiosperm order, Cornales.

It bears simple leaves with long petioles and ovate, cordate-based laminae which are serrate with prominent right-angled teeth (Fig. 2A). Inflorescences consist of globose terminal heads with a pair of showy bracts positioned some distance below on the long peduncle. The bracts are similar in size to the leaves, but thinner and less uniformly serrate, and fall away soon after anthesis. Nearly all flowers in the head are staminate, but a single bisexual flower, upon fertilization, develops into the fruit. The resulting elliptical fruit is borne on an odd looking structure representing the remnant of the flowering head with scars of the shed staminate flowers, terminating a substantial peduncle (Fig. 3A). The fruit stone is woody, composed of fibers, and has several radially arranged locules (Fig. 3C), each with a tongue shaped, elongate, germination valve that hinges open from the base.

The fossil record of Davidia extends at least back at least 58 million years, and is well documented by fruits and leaves of Davidia antiqua (Newberry) Manch. from the Paleocene of North America. The leaf fossils (Fig. 2B-D) were initially considered to represent Viburnum, because some species of that modern genus (e.g. V. dentatum, V. ellipticum, V. lantana), have leaves that are almost identical in the pattern of venation and serration. However, the fossils are distinguished from Viburnum in having long petioles, nearly as long as the lamina, as those of extant Davidia.
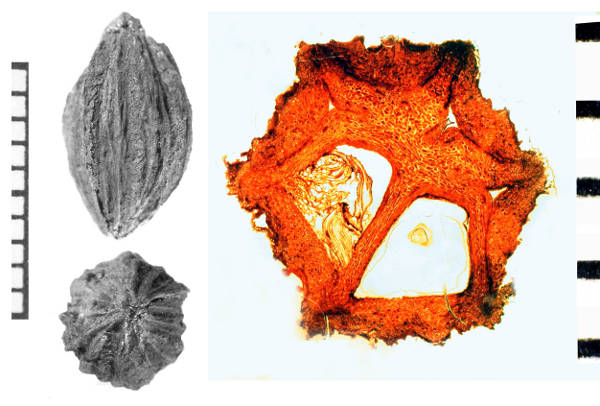
The associated fossil fruits of Davidia antiqua (Fig. 2D, E) correspond to the modern genus in being borne singly on a globose head with a long petiole that shows scars where the two bracts detached (Fig. 3B). Anatomically preserved fruits show the diagnostic anatomy, with vascular strands dispersed through the septae, rather than running along the central axis, and have 6-8 radially arranged single-seeded locules, each with a corresponding dorsal germination valve (Fig. 3D). Because the North American fossils are so similar to the living representative, Davidia deserves the title, Living Fossil. There are, however, a few differences which indicate the Paleocene fossil represents an extinct species.
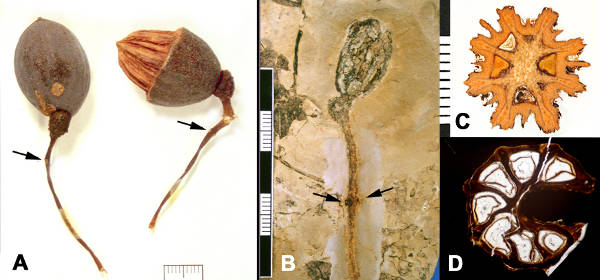
Fruits of modern Davidia involucrata are larger than those of the Paleocene fossil (25-35 mm vs 12-21 mm). Additionally, the surface sculpture of the stones in the extant species is more complicated, with a rougher texture (Fig. 3A, right) and a ridge over the midline of each locule (Fig. 3C) as well as the more prominent ridges situated between the locules; the Paleocene fossils lack the middle ridges (Fig. 3D).
Stratigraphic range and Geographic history.
The fossil record indicates that Davidia had a broader geographic range in the past than it does today (Fig. 4). The oldest probable occurrences of Davidia are permineralized fruits from the Upper Cretaceous (Campanian) Horseshoe Canyon Formation of Dinosaur Provincial Park near Drumheller, Alberta, Canada (Serbet et. al 2004; Fig. 5). Those fruits are smaller than those of D. antiqua and have fewer locules, but otherwise conform in morphology to the extant genus, although other features of the plant such as leaves and inflorescence structure remain unknown. In North America, the genus is represented also by leaves from the Early Eocene [leaves from Republic, Washington, published as Tsukada davidiifolia Wolfe & Wehr (1977)], and by permineralized fruits from the Late Eocene of the John Day Formation, Oregon (Manchester & McIntosh 2007). The genus is unknown from the European fossil record, though it would be easily recognized in the numerous leaf and carpofloras preserved there. Likely leaves and fruits are known as impressions from the early Paleocene of Far Eastern Russia (Manchester 2002). Leaves assigned to Davidia palaeoinvolucrata Pavlyutkin and associated fruits, assigned to Davidia nezhinoensis Pavlyutkin were described from the late Early Miocene of Primorye (Pavlyutkin, 2009). Although the identification appears likely to be correct, the length of the petiole, necessary to confirm that the leaves are Davidia and not Viburnum, was not described or illustrated for D. palaeoinvolucrata. In Japan, fruits and leaves indistinguishable from extant D. involucrata occur in the Pliocene to Pleistocene of Honshu (Kokawa, 1965; Ozaki, 1984; Tsukagoshi et al., 1997).
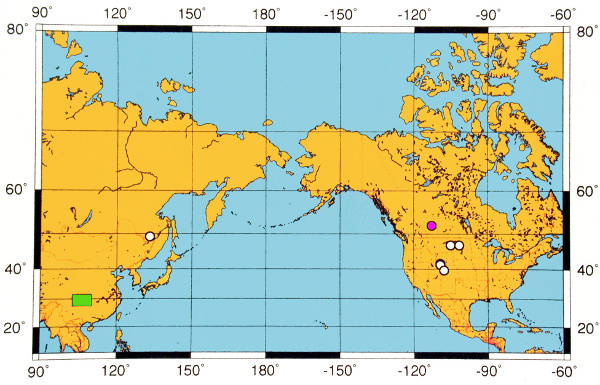
The fossil record suggests that Davidia traversed the Bering Land Bridge during the late Cretaceous or early Tertiary, allowing it to become established both in North America and eastern Asia by the Paleocene. This biogeographic pattern is similar to that of the related extinct genus Amersinia/Beringiaphyllum (Manchester et al 1999), and is distinct from that of other cornalean genera Alangium, Cornus, Mastixia and Nyssa, which successfully populated Europe as well as North America during the early Tertiary and appear likely to have traversed the North Atlantic land bridge. The restricted modern native range of Davidia in central China is the result of extinctions that obliterated the North American species, apparently at the end of the Eocene, and a reduction of once broader Asian range. Davidia is among more than 50 wood of seed gnera now endemic to eastern Asia, for which a significant fossil record has been documented (Manchester et al. 2009).
Contribution by Steven R. Manchester
Florida Museum of Natural History
Cited literature:
- Brown RW 1962. Paleocene floras of the Rocky Mountains and Great Plains. US. Geological Survey Professional Paper 375: 1–119, pls. 1–69.
- Kokawa S. 1965. Fossil endocarp of Davidia in Japan. Journal of Biology, Osaka City University 16: 45–51, 4 pls.
- Manchester SR. 2002. Leaves and fruits of Davidia (Cornales) from the Paleocene of North America. Systematic Botany 27(2): 368-382 + cover photo.
- Manchester SR, and McIntosh WC 2007. Late Eocene silicified fruits and seeds from the John Day Formation near Post, Oregon. PaleoBios 27(1):7–17.
- Manchester SR, Crane PR, Golovneva L. 1999. An extinct genus with affinities to extant Davidia and Camptotheca (Cornales) from the Paleocene of North merica and Eastern Asia. International Journal of Plant Science 160: 188-207.
- Mancheste, SR, Chen ZD, Lu AM, Uemura K. 2009 Eastern Asian endemic seed plant genera and their paleogeographic history throughout the Northern Hemisphere. Journal of Systematics and Evolution 47 (1): 1–42.
- Serbet R, Manchester SR, Aulenbach K, Braman D. 2004 Nyssaceae among the dinosaurs: anatomically preserved fruits from the Upper Cretaceous Horseshoe Canyon Formation, Drumheller, Alberta, Canada. Botany 2004 Conference, Snowbird, UT.
- Sun Ji-Fan and Huang Shuang-Quan. 2011. White Bracts of the Dove Tree (Davidia involucrata): Umbrella and Pollinator Lure? Arnoldia 68 (3): 2-10.
- Ozaki K. 1984. Two new fossil species of Fortunearia and Davidia from the upper Motojuku (Kabutoiwa) Formation in Central Japan. Memoirs of the Institute of Field Education, Yokohama National University 2: 1–8.
- Tsukagoshi M, Ono Y, Hashimoto T. 1997. Fossil endocarp of Davidia from the Early Pleistocene sediments of the Tokai Group in Gifu Prefecture, central Japan. Bulletin of the Osaka Museum of Natural History 51: 13–23.
- Wolfe JA, Wehr WC. 1987. Middle Eocene dicotyledonous plants from Republic, Northeastern Washington. U. S. Geological Survey Bulletin 1597: 1–25.
- Ying Tsun-Shen, Zhang Yu-Long, Boufford DE. 1993. The endemic genera of seed plants of China. Beijing: Science Press.Crane PR 1985. Phylogenetic analyses of seed plants and the origin of angiosperms. Annals of the Missouri Botanical Garden 72: 716-793.